クリスタルバイオレット(クリバイ)とエチルバイオレット(エチバイ)の分子構造です。両方ともtriphenylmethane色素の仲間です。
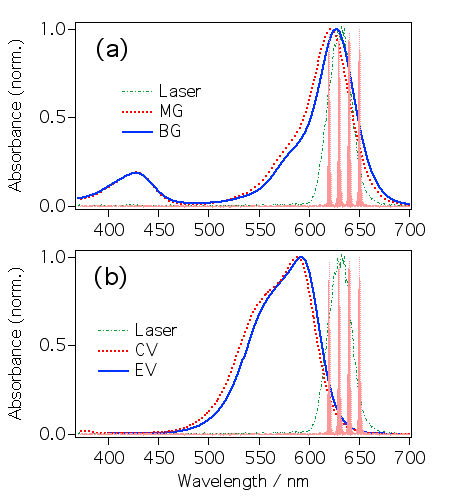
TPM色素の吸収スペクトルの比較です。マラグリ、ブリグリの吸収スペクトルには430nm付近に吸収があるのに、クリバイ、エチバイにはありません。これはどうしてでしょうか?クリバイ、エチバイの分子構造はD3という高い対称性を持ちます。その結果励起状態が2重に縮退しています。クリバイ、エチバイの吸収スペクトルの短波長側の肩は対称性の低下により縮退が若干解けているので現れると言われています。これに対し、マラグリ、ブリグリはC2と対称性が低くなり縮退が解けて430nm付近に新しい吸収が現れるのです。マラグリ、ブリグリの吸収スペクトルの短波長側の小さな肩は振動構造でしょう。
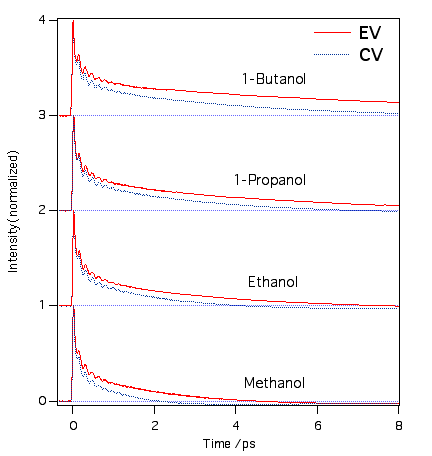
クリバイとエチバイもマラグリやブリグリのようにアルコール溶媒の炭素鎖を長くすると励起状態の寿命が延びていきます。これは励起状態の寿命が溶媒の粘度に依存していることを現しています。triphenylmethane色素に共通の性質です。エチバイのほうがクリバイよりも分子サイズが少し大きいので寿命がちょびっと長くなります。水にも溶けるのですが、アグリゲートしてしまうので測定できませんでした。
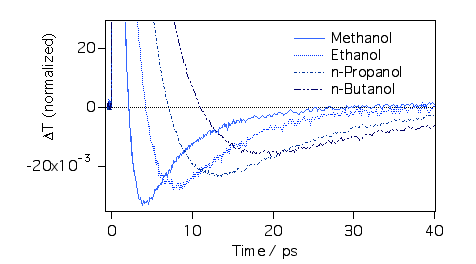
クリバイとエチバイにははっきりと負の信号が現れます。負の信号の寿命も粘度に依存し、その寿命はねじれた励起状態の寿命より2倍以上長いことがわかりました。よってこれは基底状態のホットなねじれ分子の過渡吸収に帰属しました。
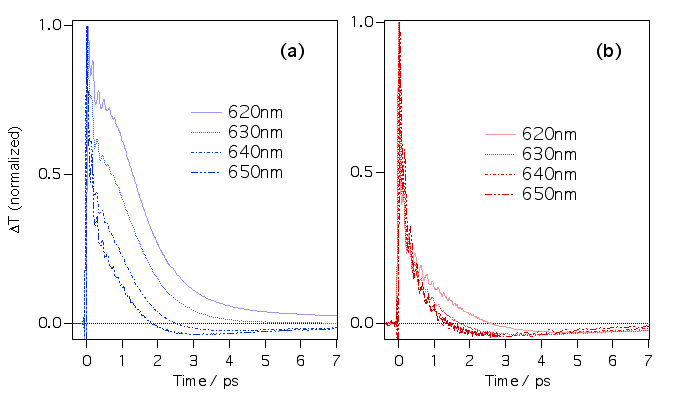
(a)マラグリ/メタノールの信号、(b)クリバイ/メタノールの信号を波長分解したものです。マラグリの場合、長波長側にのみ負の信号が現れ、短波長側では正の信号に変わっていることがわかります。このことも負の信号が基底状態のホットなねじれ分子の過渡吸収であるとこを裏付けています。クリバイの場合、もともと吸収スペクトルの長波長端を励起しているのでそのような効果は現れません。
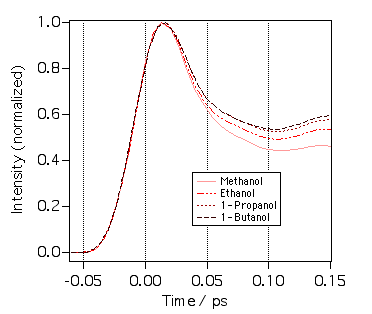
上の図はクリバイのPP信号の比較です。驚いたことに溶媒依存性は非常に速い時間領域から現れています。50フェムト秒の時点でメタノールに比べてブタノールの減衰がすでに遅くなっているのがわかります。これにはちょっと驚きました。溶媒和の慣性項がアルコールの炭素鎖長に依存するということなのでしょうか?別の言葉でいうと、ベンゼン環が最初の溶媒分子と衝突した瞬間に衝突した相手がどんな奴か認識しているということでしょう。トラックなのか軽自動車なのかぐらいはすぐわかるということですな。
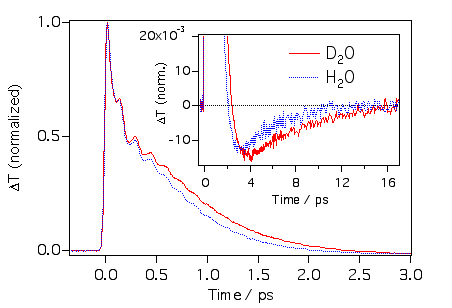
もしかして水と重水でも違いが出るかもしれないと思ったので、比較してみました。クリバイ、エチバイは水に溶かすとアグリゲートしてしまう(アグってしまう)のでマラグリで実験をしてみました。予想通り違いがでましたが、水と重水では粘度が違う(1.0と1.2cPぐらい)のでその効果が出ていると思われます。ちなみに一番速いところのダイナミクスには違いが見られませんでした。これは水素結合のlibrationは慣性項に寄与しないということなのでしょうか?
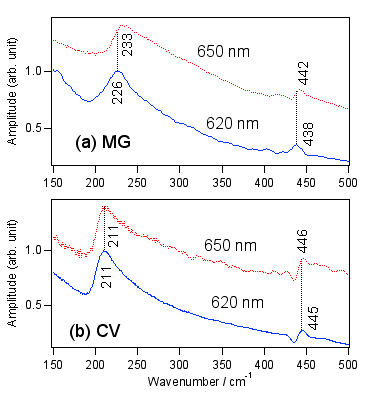
TPM色素の信号にはビートが出ているのでフーリエ変換すると分子振動の波数がわかります。210-230cm-1の振動はTPM色素特有の中心炭素周りのブリージングモードだと言われています。
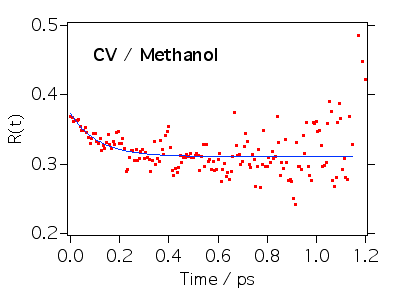
クリバイの異方性は100フェムト秒程度で減衰することがわかりました。クリバイは吸収スペクトルの長波長端を励起しているので誘導放出(蛍光)の寄与が大きいと考えられます。TPM色素は励起状態でフェニル基がねじれると遷移双極子の大きさが小さくなると言われています。この結果は遷移双極子は小さくなるだけではなくてその向きも変わっているということを表しています。またクリバイの励起状態は縮退しており、その状態の遷移双極子同士は直交しているので縮退した状態間の熱平衡化過程が見えている可能性もあります。
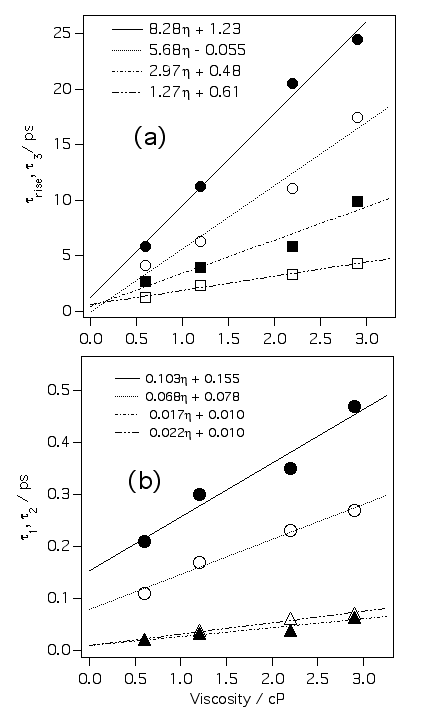
クリバイ、エチバイの信号を多指数関数でフィッティングしてやり、その各成分の粘度依存性をプロットすると上の図ようになります。△、○、□はクリバイ、▲、●、■はエチバイの各成分です。おもしろいことに遅い成分ほど粘度に対する傾きが急になっています。我々はこれは溶質周りの溶媒和殻の階層構造の現れであると考えています。ねじれ始めたフェニル基はまず第一溶媒和圏の数個の溶媒分子と衝突します。時間とともにフェニル基に衝突する分子の数は増え、衝突した分子は別の溶媒和分子に衝突します。このように時間とともに衝突する分子数が増えるため、粘度依存性が時間に依存すると考えているわけです。
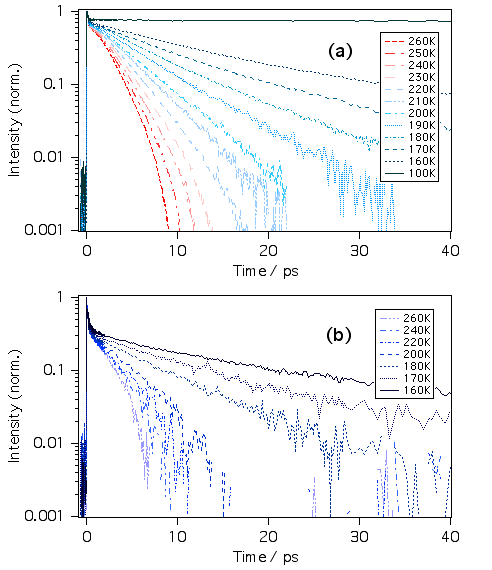
さて、溶媒和殻の階層構造をより詳しく調べるため、温度依存性も実験してみました。(a)はマラグリ、(b)はクリバイのエタノール溶液です。温度下降とともに減衰が遅くなるのがわかります。
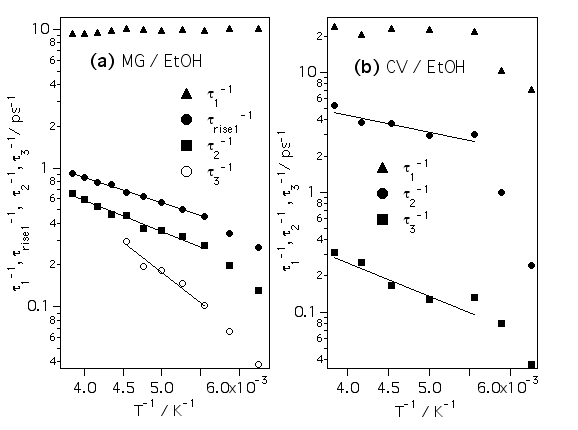
アーレニウスプロットをしてみたところ、遅い成分ほど活性化エネルギーが大きいことがわかりました。TPM色素のフェニル基の回転は活性化エネルギーがゼロだと言われているので、この活性化エネルギーは溶媒由来であると考えられます。これも溶媒和殻の階層構造を考えると、時間が経つほど衝突する溶媒分子の数が増えるので、活性化エネルギーも大きくなると結論できます。
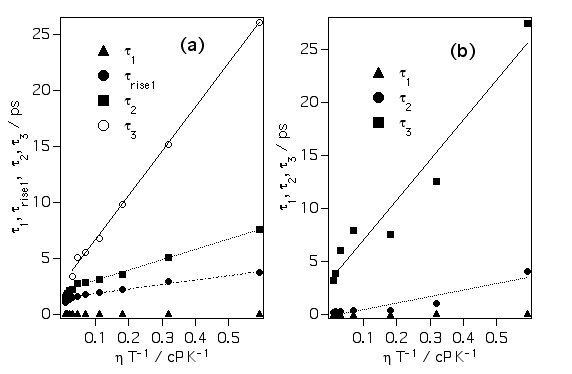
さらに各寿命を粘度/温度に対してプロットするとおもしろいことがわかります。ストークス・アインシュタイン・デバイ理論によると、回転緩和時間は粘度/温度に対して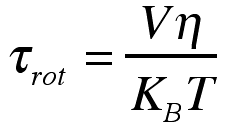
の関係があります。ここでVは有効体積です。上の図を見るとわかるように遅い成分ほど傾きが大きくなっている、つまり有効体積が増えていることがわかります。実際のTPM色素の体積は変化しないのに見掛けの体積が変化しているということは、時間とともに回転運動に関わる溶媒分子の数が増えるということで説明でき、これも溶媒和殻の階層構造を裏付ける結果と言えます。