まあ、いきなり蛋白質について調べるのは敷き居が高いので、今回は糖水溶液の濃度と粘度の関係を調べてみたわけです。
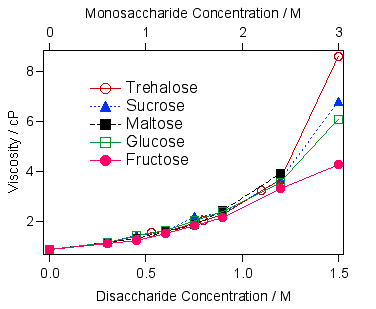
図1 糖水溶液の濃度と粘度の関係をオストワルド粘度計で測ると、このようになります(25℃)。濃度の低いところでは、ほどんど糖による差は現れません。ところが濃度が1.5 Mになると、トレハロースの粘度が一番高くなります。トレハロースは二糖類なので、単糖類については二倍の濃度と比較しているのですが、それでもトレハロースの粘度の方が高いのがわかります。粘度は糖濃度で非線形に増加していくので、濃度の高いところで糖は会合体を形成しているのではないか?と考えました。そして、トレハロースの会合能が一番大きいと解釈しました。

図2 そこで、マラカイトグリーンをプローブ分子としてミクロスコピックな粘度を見てみることにしました。マラカイトグリーンの第一電子励起状態の寿命は溶媒のミクロな粘度に依存することが知られています。(関連ページ:その1 その2)
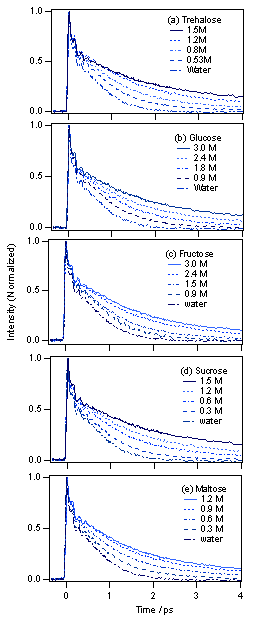
図3 マラカイトグリーンの励起状態寿命は糖濃度上昇とともに長くなっていくことがわかります。

図4 (a) 濃度の低いところでマラカイトグリーンの減衰曲線を対数プロットしてみると、ピコ秒領域では直線的になり減衰が指数関数的であることがわかります。(b) ところが、濃度が高くなると減衰は曲線的になり、二成分の指数関数でないとフィッティングできなくなることがわかります。残差1は1つのピコ秒成分のみでフィッティングした場合、残差2は2つに増やした場合のもので、フィッティングの精度がよくなっていることがわかります。この結果は溶液の不均一性が大きくなっていることを表しており、トレハロース会合体の近傍では粘度が非常に高いのに対し、遠いところではそれほど粘度は高くなってないことを表していると考えられます。
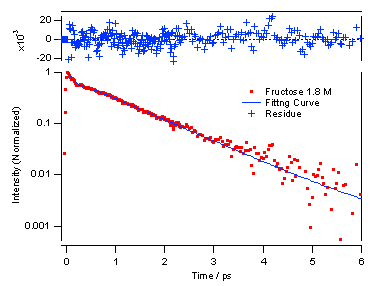
図5 単糖類のフルクトースについては、濃度をけっこう上げても指数関数的に減衰しており、溶液が均一であることがわかります。これは二糖類であるトレハロースの方が会合して不均一になりやすいということを示しています。
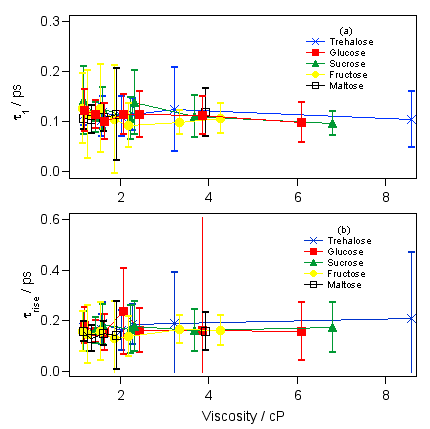
図6 マラカイトグリーンの減衰曲線は全部で3つの減衰成分と1つのライズ成分でフィッティングできますが、そのうちの300フェムト秒以下の2つの超高速成分の時定数は糖濃度に依存しません。この時間領域では糖の影響はほとんどなく、バルクの水溶液のダイナミクスと大差ないことがわかります。
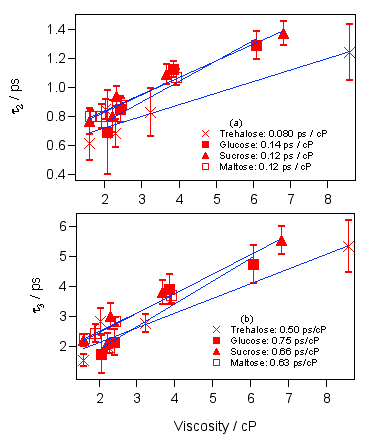
図7 ところが、0.5ピコ秒より遅い成分になると、はっきりと粘度依存性が出てくることがわかります。おもしろいことにどちらのピコ秒成分についてもトレハロースの濃度依存性の傾きが一番小さいことがわかります。このことは、トレハロースは高率よく会合するので、巨視的な粘度は大きくなりますが、ミクロな流動性はそれほど失われず、マラカイトグリーンのベンゼン環は比較的自由に動くことができるということを示していると考えられます。